水稻制種轉(zhuǎn)育是啥_如何獲得水稻轉(zhuǎn)基因植株?
我國的水稻育種經(jīng)歷了矮化育種、雜種優(yōu)勢利用和綠色超級稻培育3次飛躍,其間伴隨矮化育種(第一次綠色革命)、三系雜交稻培育、二系雜交稻培育、亞種間雜種優(yōu)勢利用、理想株型育種和綠色超級稻培育等6個重要歷程。育種目標從唯產(chǎn)量是舉到高抗、優(yōu)質(zhì)和高產(chǎn)并重,育種理念從高產(chǎn)優(yōu)質(zhì)逐步提升為“少投入,多產(chǎn)出,保護環(huán)境”。水稻功能基因組研究為第二次綠色革命準備了大量的有重要利用價值的基因,水稻育種正邁向設(shè)計育種的新時代。基因組選擇技術(shù)和轉(zhuǎn)基因技術(shù)將為培育“少打農(nóng)藥,少施化肥,節(jié)水抗旱,優(yōu)質(zhì)高產(chǎn)” 綠色超級稻保駕護航。
本文對我國水稻遺傳育種的發(fā)展歷程進行了概括,指出了各種育種方法和育種技術(shù)的優(yōu)缺點,系統(tǒng)介紹了水稻細胞質(zhì)雄性不育和光溫敏雄性核不育以及秈粳雜種不育的分子機制的研究進展,綜述了水稻株型、穗型、粒形和養(yǎng)分高效利用相關(guān)的重要功能基因,闡明了產(chǎn)量與開花期聯(lián)動的關(guān)系,凸顯了我國水稻基礎(chǔ)研究在國際上的重要地位。特別指出,近年來,我國水稻生產(chǎn)方式發(fā)生了或正在發(fā)生巨大變革,育種理念也要與時俱進。未來,雜交育種技術(shù)要與現(xiàn)代育種技術(shù)緊密結(jié)合,選育水稻品種不僅要滿足市場需求,而且更要具備綠色健康的特點,同時還要適應(yīng)新耕作制度和新耕作方法。
在水稻種植歷史中,無論是常規(guī)地方品種、現(xiàn)代品種還是雜交組合,產(chǎn)量一直是重要考量目標之一。特別是20世紀中期以來,我國農(nóng)業(yè)生產(chǎn)力低下,為了解決糧食短缺問題,我國育種家唯產(chǎn)量是舉,培育出耐大肥大水的品種,依靠大量使用農(nóng)藥化肥,雖然解決了燃眉之急,但也給我國的生態(tài)環(huán)境造成了破壞,這種生產(chǎn)模式不具有可持續(xù)性。21世紀以來,人們生活水平顯著提高,我國的水稻生產(chǎn)漸漸地發(fā)生了重大變化,育種目標也從單一高產(chǎn)轉(zhuǎn)向優(yōu)質(zhì)高抗高產(chǎn)等復合性狀目標,綠色農(nóng)業(yè)指日可待。
1 水稻遺傳育種6個重要歷程
1.1 水稻第一次“綠色革命”
水稻地方品種基本是高稈類型,耐肥力差,容易倒伏,導致穩(wěn)產(chǎn)問題。因此,發(fā)掘矮稈種質(zhì)資源,培育矮稈品種變得十分重要。1956年,我國育種家黃耀祥先生以廣西農(nóng)家品種“矮仔占”為材料,選育出“矮仔占4號”,并與高稈品種“廣場13”雜交,培育出第一個矮稈秈稻品種“廣場矮”。1966年,國際水稻研究所(IRRI)利用我國臺灣省地方品種低腳烏尖(Dee-geo-woo-gen)與皮泰(Peta)雜交,育成了半矮化的品種IR8,創(chuàng)造了當時的高產(chǎn)奇跡。我國的矮化育種比國際上整整提前了10年。“廣場矮”的培育以及IR8的引進推動了我國水稻育種進入第一次“綠色革命”時代,水稻畝產(chǎn)從50年代的164千克提高到70年代初的238千克,水稻單產(chǎn)提高了將近45%,實現(xiàn)了水稻產(chǎn)量的第一次飛躍。實際上矮稈品種單株產(chǎn)量略微下降,但是由于其耐密植和抗倒伏,群體產(chǎn)量大幅度提升;同時矮稈品種的耐肥能力強,氮素吸收利用效率低,大量氮肥的施用在提高產(chǎn)量的同時使土壤和環(huán)境受到破壞。
1.2 核質(zhì)互作雄性不育系的培育和水稻三系雜種優(yōu)勢利用
雜種優(yōu)勢是指一個物種的不同品種或者物種間的雜交后代的生物量、發(fā)育速度和產(chǎn)量的表型值優(yōu)于兩個親本的現(xiàn)象。我國稻作科學的奠基人丁穎先生曾用人工辦法給水稻“去雄”,但實際效果不佳。1964年,湖南安江農(nóng)校的袁隆平先生開始雜交水稻育種研究,并提出雜種優(yōu)勢利用的設(shè)想。1970年,袁隆平先生和他的助手李必湖等人在海南三亞發(fā)現(xiàn)了花粉敗育的野生稻,花粉敗育是由不育細胞質(zhì)產(chǎn)生(CMS- WA),為雜交水稻雄性不育系的選育打開了突破口。1971年,雜交水稻課題被列為全國協(xié)作項目,野敗材料被分發(fā)到國內(nèi)水稻科研單位。1973年10月,在蘇州召開的水稻科研會議上,袁隆平先生發(fā)表《利用“野敗”選育“三系”進展》的論文,標志中國秈型雜交水稻三系(不育系、保持系和恢復系)配套成功。同時,江西省萍鄉(xiāng)市農(nóng)業(yè)科學研究所的顏龍安先生利用“野敗”育成不育系珍汕97A。1981年,福建省三明市農(nóng)業(yè)科學研究所的謝華安先生育成具有抗病性強、配合力高、米質(zhì)優(yōu)良的恢復系明恢63。明恢63與珍汕97A配制的強優(yōu)勢組合汕優(yōu)63產(chǎn)量高,且具有抗病、耐低磷鉀、耐高低溫、米質(zhì)較好、適應(yīng)性廣等特點,累計推廣6287.7萬公頃。三系雜交稻平均產(chǎn)量比一般普通良種增產(chǎn)20%左右,畝產(chǎn)能達到430公斤。
CMS-WA是細胞質(zhì)基因和核基因互作導致花粉敗育類型,屬于孢子體雄性不育。可以通過回交的方法保留細胞質(zhì)基因組而交換核基因組,達到培育不育系的目的。除CMS-WA外,我國水稻育種學家還創(chuàng)制出不同細胞質(zhì)來源的核質(zhì)互作雄性不育系。四川農(nóng)業(yè)大學的周開達先生等用西非品種岡比亞卡與朝陽1號、雅安早等雜交和回交,育成岡型不育系岡12朝陽1號A和岡22雅安早A;同時,周開達先生等從Dissi D52/37//矮腳南特群體中選出不育株,育成D型不育系意大利A。意大利A與珍汕97B中變異株雜交,育成D汕A。湖南省農(nóng)業(yè)科學院利用地理遠距離品種間雜交組合,育成印尼水田型不育系。岡型、D型和印尼水田型不育特性以及恢、保關(guān)系與野敗相似。不同育種單位育成了很多孢子體雄性不育類型的不育系,如四川省農(nóng)業(yè)科學院的K型,其胞質(zhì)不育基因來自于粳稻,恢、保關(guān)系與野敗相同;安徽省廣德縣農(nóng)業(yè)科學研究所從江西省引進的矮桿野生稻中發(fā)現(xiàn)一株雄性不育株,命名“矮敗”,通過核置換育成協(xié)青早A,恢、保關(guān)系與野敗相同,K型和矮敗型不育系屬于孢子體不育。1972年,武漢大學朱英國先生等以紅芒野生稻為母本與蓮塘早雜交,選育出紅蓮型細胞質(zhì)雄性不育系(HL- CMS),為配子體不育類型。包臺型(BT)不育系臺中65A引入我國,并經(jīng)湖南省農(nóng)業(yè)科學院轉(zhuǎn)育成BT型黎明A,與C系統(tǒng)恢復系的配組,使得BT型不育系成為我國粳稻雜種優(yōu)勢利用的主要不育系類型。BT型不育系花粉敗育屬于配子體不育,不育性沒有CMS-WA穩(wěn)定,高溫易自交結(jié)實,雜交制種種子不純。滇型不育系是云南省培育出的適應(yīng)當?shù)馗吆0螝夂颦h(huán)境的粳稻CMS,屬于配子體不育類型。HL-CMS的花粉敗育特征為圓敗型,而BT- CMS和滇型為染敗型。
1.3 光溫敏雄性核不育系的培育和水稻兩系雜種優(yōu)勢利用
1973年,湖北沙湖原種場農(nóng)技員石明松先生發(fā)現(xiàn)水稻農(nóng)墾58的光敏核不育(PGMS)株,并育成了首個光敏核不育系農(nóng)墾58S。農(nóng)墾58S在長日高溫條件下表現(xiàn)為雄性不育,作為不育系用于雜交水稻制種;在短日低溫條件下可育,用于不育系的繁種。PGMS的敏感階段為幼穗發(fā)育時期,主要是從二次枝梗分化到花粉母細胞形成時期,在長日照條件下絨氈層提前降解,缺乏營養(yǎng)供給導致小孢子敗育。溫度和光照長度在農(nóng)墾58S中具有補償效應(yīng),高溫可以降低臨界日照長度,而低溫要提高臨界日照長度。利用農(nóng)墾58S光敏不育的特性,突破三系配套恢保關(guān)系束縛,開創(chuàng)了“二系”雜交水稻育種的新階段。另外一類種質(zhì)例如安農(nóng)S-1、衡農(nóng)S-1和5460S,稱為水稻溫敏核不育(TGMS),溫度變化可以導致育性的轉(zhuǎn)換,高溫不育,低溫可育,而光周期長短對育性轉(zhuǎn)換沒有影響。TGMS的誘導階段在花粉母細胞形成和減數(shù)分裂時期,小孢子母細胞不能完成減數(shù)分裂,敗育的花粉都呈現(xiàn)出皺縮的形態(tài)。兩系雜交稻由于沖破了恢保關(guān)系的束縛,親本間的遺傳差異變大,兩系雜交稻平均產(chǎn)量比三系雜交稻具有較大的提高,代表性的品種如兩優(yōu)培九大面積種植能達到畝產(chǎn)630公斤,比三系對照汕優(yōu)63增產(chǎn)約10%左右。
1.4 秈粳亞種間雜種優(yōu)勢的利用
秈稻和粳稻亞種之間具有更豐富的遺傳多樣性,雜交組合比秈秈組合具有更強的雜種優(yōu)勢。但是,秈粳雜交種F1不育(或部分可育)限制了秈粳雜種優(yōu)勢的利用。廣親和基因的發(fā)現(xiàn)為秈粳亞種間雜種優(yōu)勢利用奠定了理論基礎(chǔ)。利用部分粳稻血緣培育的雜交組合例如兩優(yōu)培九、協(xié)優(yōu)9308等表現(xiàn)出很強的雜種優(yōu)勢。直到21世紀以來,利用粳稻不育系與秈粳中間型廣親和恢復系配組配制出秈粳亞種間的雜交品種,如“甬優(yōu)系列”和“春優(yōu)系列”組合。這些雜交組合在生產(chǎn)上表現(xiàn)出更強的產(chǎn)量優(yōu)勢。
1.5 理想株型育種
理想株型是由澳大利亞科學家CM. Donald博士提出,指農(nóng)作物個體間競爭最小的株型,使每個植株最大限度地獲取光照和營養(yǎng),從而提高群體的收獲指數(shù)。日本栽培學家松島省三最早提出高產(chǎn)水稻應(yīng)該具備“多穗、矮桿、短穗,頂部2、3葉片要短厚直立”的特性。Khush提出少蘗、大穗、莖稈粗壯、株高100~110 cm、葉厚直立、根系發(fā)達、晚熟、收獲指數(shù)高和生產(chǎn)潛力大等特征。我國育種家楊守仁先生等提出高產(chǎn)水稻指標:半矮桿、穗大且直立、分蘗中等,耐肥抗倒、生物量大、谷草比高。袁隆平先生認為超高產(chǎn)雜交水稻在形態(tài)上主要特點是上部3片功能葉要長、直、窄、凹、厚,葉面積較大,并且可以兩面受光而互不遮蔽,提出庫大源足是高產(chǎn)的前提,新株型特征和雜種優(yōu)勢利用相結(jié)合是實現(xiàn)超高產(chǎn)水稻育種技術(shù)路線。2010年,李家洋先生克隆出理想株型基因IPA1。IPA1植株株型緊湊,莖稈挺直,雖然分蘗能力不強,但成穗率高、穗大、產(chǎn)量潛力大。IPA1的克隆促進了理想株型的育種,培育的理想株型新品種已經(jīng)表現(xiàn)出巨大的增產(chǎn)潛力。
1.6 第二次綠色革命理念及綠色超級稻品種選育
1999年12月14日,西北農(nóng)林科技大學的李振聲先生、華中農(nóng)業(yè)大學的張啟發(fā)先生和中國農(nóng)業(yè)科學院作物科學研究所賈繼增先生在杭州召開的“農(nóng)作物資源核心種質(zhì)構(gòu)建、重要新基因發(fā)掘與有效利用”973項目年會中提出了第2次綠色革命的10字口號,“少投入,多產(chǎn)出,保護環(huán)境”,并提出為綠色革命準備基因資源。國家973項目“作物高效利用氮磷養(yǎng)分的分子機理”、“主要糧食作物重大病害控制的基礎(chǔ)研究”和“害蟲爆發(fā)成災的遺傳與行為機理”以及農(nóng)業(yè)部948項目“參與全球水稻分子育種計劃研究”等,推動了作物營養(yǎng)高效利用和對逆境抗性基因的發(fā)掘。2005年和2007年,張啟發(fā)先生先后兩次撰文,提出培育綠色超級稻的構(gòu)想,主要內(nèi)容包括“少打農(nóng)藥,少施化肥,節(jié)水抗旱,優(yōu)質(zhì)高產(chǎn)”,將第二次綠色革命的基本理念貫穿始終。經(jīng)過10年的努力,我國選育出一批綠色超級稻品種,全國累計推廣約9000萬畝。
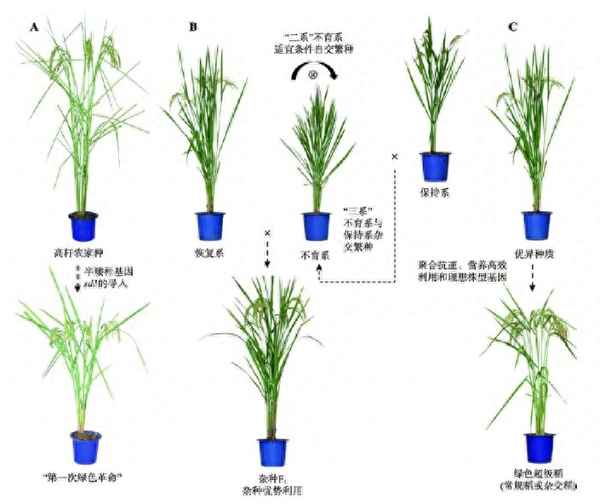
圖1 水稻遺傳育種經(jīng)歷的3次飛躍。A:半矮桿基因sd1利用以及半矮稈水稻品種培育促成“第一次綠色革命”;B:不育系和恢復系配制雜種促進雜種優(yōu)勢的利用;C:抗逆、養(yǎng)分高效利用和理想株型等有利基因的發(fā)掘促進綠色超級稻的培育。
水稻遺傳育種的歷程就是一個育種理念變遷的過程。我國水稻遺傳育種經(jīng)歷了3次大的飛躍(圖1),每次飛躍都離不開重要基因資源的發(fā)掘和利用。矮稈基因?qū)е隆暗谝淮尉G色革命”,解決了水稻耐肥和抗倒伏的問題;核質(zhì)互作不育系和光溫敏核不育系的培育,促成了雜種優(yōu)勢的利用。第三次飛躍以理想株型塑造為主要技術(shù)路線,以綠色超級稻育種為目標,選育高產(chǎn)優(yōu)質(zhì)健康新品種/組合,實現(xiàn)第二次綠色革命。
2 水稻遺傳育種技術(shù)的變遷
2.1 常規(guī)育種
常規(guī)育種包括選擇育種、有性雜交育種、物理以及化學誘變育種、離體組織培養(yǎng)育種和細胞雜交育種。常規(guī)育種的過程主要是選擇合適的親本,得到分離的群體,利用該育種方法,根據(jù)表型從群體后代中選擇達到所設(shè)定育種目標的個體。這種方法對高產(chǎn)育種效率比較高,但是對稻米品質(zhì)和非生物逆境的改良效率較低。選擇育種是從自然變異中選擇優(yōu)良變異,但是自然變異發(fā)生頻率低,有價值變異少,育種效率低。誘變育種是通過物理化學處理,增加誘變頻率,從大量突變中選擇有利突變,由于突變往往是有害的,因此育種效率也低。利用水稻花藥培養(yǎng)再生植株,單倍體自然加倍,基因組純合快,能大量縮短育種歷程,但是花藥培養(yǎng)嚴重依賴于基因型,特別是秈稻的花藥培養(yǎng)難度較大,因此花藥培養(yǎng)育種也受到限制,只有少數(shù)單位開展。有性雜交育種是利用不同親本材料雜交,再通過自交或測交,產(chǎn)生大量的具有豐富表型變異的后代群體,從中選擇優(yōu)良表型的單株。雜交育種充分發(fā)揮基因重組的作用,只要親本間互補性強,雜交育種效率一般比較高,并且很可能育成全新的骨干品種。因此,雜交育種是最主流的水稻育種方法,得到廣泛應(yīng)用。
2.2 分子標記輔助選擇育種和基因組育種
近30年來水稻功能基因組學的成果為輔助選擇育種提供了一系列的功能分子標記。SNP芯片是全基因組選擇育種的有效工具,水稻60K SNP芯片的開發(fā)和應(yīng)用為大規(guī)模的基因型鑒定提供了便捷的方法。在這些標記輔助選擇下,通過回交實現(xiàn)目標性狀的定向改良。2017年,縮短作物生長周期的快速育種方法誕生,該方法可以使春小麥和豌豆等實現(xiàn)一年種植6代,油菜一年可以種植4代,加速了育種進程。水稻是短日照植物,結(jié)合基因組選擇育種和快速育種方法,可充分發(fā)揮定向改良的效率。定向改良必須知道哪些基因(等位基因)具有控制有利農(nóng)藝性狀和生物學性狀的功能。在育種過程中,水稻材料大多在正常生產(chǎn)條件下種植,抗性性狀如抗生物逆境和非生物逆境很難通過田間目測加以選擇,而標記輔助選擇在苗期就可以進行。因此,利用標記輔助育種和基因組育種定向改良抗逆等性狀更有現(xiàn)實意義。隨著功能基因的不斷挖掘和基因調(diào)控網(wǎng)絡(luò)的建立,全基因組范圍的設(shè)計育種將有更廣闊的天地。
2.3 轉(zhuǎn)基因育種與基因編輯育種
轉(zhuǎn)基因育種是指通過轉(zhuǎn)基因的方法,導入外源的基因,達到性狀改良的目標,從而培育出新品種。傳統(tǒng)育種只能依靠品種或者種之間的雜交實現(xiàn)重組,選育出具有優(yōu)良性狀的品種。而轉(zhuǎn)基因育種可以實現(xiàn)跨物種的基因交流,對目標性狀改良的針對性強,提高育種效率。蘇云金芽胞桿菌的Bt毒蛋白基因是目前世界上公認的高效抗蟲基因之一,通過轉(zhuǎn)基因的方法將其導入水稻,可以有效提高水稻的抗蟲特性。bar基因能使植物特異性獲得對除草劑草丁膦的抗性,轉(zhuǎn)bar基因的抗除草劑水稻能獲得很好的抗除草劑效果。
近10年來,基因編輯技術(shù)的突飛猛進,特別是CRISPR/Cas9技術(shù)的應(yīng)用,基因敲除技術(shù)已經(jīng)成為常規(guī)技術(shù),基因敲入技術(shù)也產(chǎn)生了突破。因此,定向敲除不良目標基因和定向整合優(yōu)良目標基因,將大幅提高水稻定向遺傳改良效率。并且,該系統(tǒng)獲得的植株通過自交重組,容易得到不含轉(zhuǎn)基因的基因編輯品種。
圖2 3種水稻育種方法流程(以選育矮稈抗病品種為例)。A:常規(guī)育種;B:標記輔助快速育種;C:轉(zhuǎn)基因育種。
以選育矮稈抗病品種為例,3種育種方法各有千秋(圖2)。常規(guī)育種歷時長,但可以培育出全新的矮稈抗病品種;輔助選擇快速育種歷時大大縮短,定向改良矮稈抗病性狀,獲得與親本綜合性狀類似的矮稈抗病品種;而轉(zhuǎn)基因育種可以在較短的時間(2~3世代)內(nèi)獲得與受體相似的矮稈抗病品種。
3 水稻遺傳育種分子機制研究進展
水稻基因組測序的完成掀起了水稻基因功能研究的熱潮。基于自然變異的正向遺傳學策略,大量具有重要應(yīng)用價值的基因相繼被克隆,一批功能基因的分子機制得到解析,為水稻的遺傳改良提供了重要基因。
3.1 雄性不育分子機制解析
近年來,野敗型細胞質(zhì)雄性不育的分子機理得到解析。劉耀光教授課題組報道了一個野生稻線粒體中新近起源的基因WA352,與核編碼的線粒體蛋白COX11互作,共同調(diào)控水稻CMS-WA。WA352誘導雄性不育可以被兩個育性恢復基因Rf3和Rf4恢復,Rf3暫時未被克隆,Rf4編碼一個PPR蛋白,可以降低WA352的表達。BT-CMS中orf79與atp6共轉(zhuǎn)錄形成B-atp6-orf79,控制包臺型CMS。Rf1位點存在兩個緊密連鎖的恢復基因Rf1A和Rf1B,編碼PPR家族蛋白,Rf1A和Rf1B分別參與了對雙順反子mRNA B-atp6-orf79的切割和降解。與BT-CMS不同的是,HL-CMS不育系中orfH79有兩種轉(zhuǎn)錄產(chǎn)物,一種為atp6-orfH79,另一種為orfH79,并且都能翻譯成細胞毒素肽ORFH79。OrFH79蛋白能與電子傳遞鏈復合體Ⅲ的亞基P61互作,導致花粉敗育。CMS中線粒體基因突變改變了線粒體的正常狀態(tài),導致不育;而育性恢復基因編碼PPR家族蛋白,抑制或者參與對線粒體中CMS基因的mRNA加工,從而導致育性的恢復。
我國科學家在光溫敏雄性核不育的機理方面也進行了系統(tǒng)的研究。Fan等發(fā)現(xiàn)PMS1編碼一個長鏈非編碼RNA(lncRNA) PMS1T,在幼穗中表達量較高,是miR2118的作用靶標,切割產(chǎn)生21nt的phasiRNAs。在長日照條件下,農(nóng)墾58S中miR2118切割位點附近的S2突變,改變了RNA的二級結(jié)構(gòu),增強了miR2118切割效率,產(chǎn)生更多的phasiRNAs,導致不育。Ding等和Zhou等發(fā)現(xiàn)PMS3編碼一個lncRNA,分別命名為LDMAR和osa-smR5864w。LDMAR中一個G-C突變導致了RNA二級結(jié)構(gòu)的改變,并且LDMAR的啟動子區(qū)域甲基化程度升高,導致長日照條件下幼穗中表達量降低,產(chǎn)生農(nóng)墾58S中的雄性不育。
安農(nóng)S-1是第一個培育成功的溫敏不育系,受一個隱性核不育基因TMS5控制,TMS5編碼RNase ZS1,tms5一個SNP的突變導致編碼蛋白提前終止。與野生型相比,tms5中3個泛素60S核糖體蛋白UbL401、UbL402和UbL404受高溫誘導表達,蛋白質(zhì)活性實驗證明,RNase ZS1能特異切割UbL40 mRNA。安農(nóng)S-1中tms5不能對UbL40 mRNA進行切割降解,導致高溫條件下UbL40 mRNA的過度積累,從而影響細胞內(nèi)泛素平衡,引發(fā)花粉母細胞液泡化,最終導致高溫條件下花粉敗育。
3.2 雜交種F1不育的分子機制
廣親和材料的發(fā)掘以及秈粳雜種不育基因的克隆和應(yīng)用是有效解決秈粳雜種不育的根本出路。Ikehashi和Araki提出廣親和基因的遺傳模型,即廣親和的S5n等位基因型、秈稻的S5i等位基因型和粳稻的S5j等位基因型,廣親和的S5n無論是與秈稻還是粳稻雜交均可育。S5位點有3個緊密連鎖的基因—ORF3、ORF4和ORF5,其中ORF5和ORF4分別扮演了“殺手(killer)”和“幫兇(partner)”的角色,引起內(nèi)質(zhì)網(wǎng)逆境,而ORF3起到了“保鏢(protector)”的角色,對內(nèi)質(zhì)網(wǎng)逆境有清除作用。典型的秈稻基因型為ORF3+ORF4-ORF5+,典型的粳稻基因型為ORF3-ORF4+ORF5-,而廣親和品種含有ORF5n。ORF4和ORF5為孢子體型作用方式(ORF4和ORF5對所有的配子都能發(fā)揮作用),ORF3為配子體型作用方式(只對含有該基因的配子起作用)。秈粳雜種F1雌配子形成過程中,由于秈型配子ORF3+的存在,可以為其正常發(fā)育提供保護,粳型配子攜帶ORF3-不能有效防護ORF4+ORF5+的殺傷,從而導致敗育。
Sa是一個控制雜種雄配子不育基因,由兩個緊密連鎖的基因SaM和SaF組成,分別編碼泛素修飾E3連接酶和F-box蛋白。大多數(shù)的秈稻攜帶SaM+SaF+基因型,粳稻攜帶SaM-SaF-基因型,SaM-內(nèi)含子中一個SNP的改變導致剪接模式的改變,從而造成SaM-蛋白翻譯提前終止。秈梗雜種F1中存在SaM+的前提下,SaF+能與SaM-互作,導致SaM-基因型花粉敗育,稱為“兩基因/三元件”互作模型。因為SaM+多出一個自我抑制結(jié)構(gòu)域,阻止SaF和SaM+互作,SaF不能與SaM+互作。
雜種雄性不育位點qHMS7也包括兩個緊密連鎖的基因ORF2和ORF3,分別編碼核糖體失活蛋白和含線粒體信號肽的禾本科特異蛋白。滇粳優(yōu)1號攜帶有功能的ORF2DORF3D,而南方野生稻只攜帶一個無功能的ORF2N,完全缺失ORF3。ORF2D是一個毒性蛋白,以孢子體方式發(fā)揮作用;ORF3D是一個解毒蛋白,以配子體方式發(fā)揮作用。雜合(ORF2DORF3D/ORF2N-)條件下,ORF2D蛋白能殺死不攜帶ORF3D基因的花粉。因此,只有攜帶ORF2DORF3D基因型的花粉能遺傳到后代,稱為自私基因。
日本研究者發(fā)現(xiàn)兩個雜種不育位點S27和S28存在上位性互作,圖位克隆發(fā)現(xiàn)S27和S28是重復基因,編碼線粒體核糖體蛋白L27。在栽培稻臺中65中,S27有功能(T+T+)而S28無功能(TSTS),基因型為T+T+|TSTS;在展穎野生稻中,S27無功能(GSGS)而S28有功能(G+G+),基因型為GSGS|G+G+;后代中基因型為GS|TS的花粉敗育。
3.3 穗發(fā)育的分子機制
穗長、一次枝梗數(shù)、二次枝梗數(shù)和著粒密度決定每穗穎花數(shù)。Gn1a編碼細胞分裂素氧化酶基因OsCKX2,促進細胞分裂素的降解,突變后細胞分裂素得到積累,每穗穎花數(shù)增加,從而增產(chǎn)。直立密穗基因DEP1編碼G蛋白γ亞基。DEP1蛋白通過調(diào)控OsCKX2的表達影響分生組織的活性和細胞的分裂增殖,該基因突變促進細胞分裂,枝梗數(shù)增加、每穗粒數(shù)增多。FZP決定穗分枝向小穗形成的轉(zhuǎn)化,同時還抑制腋分生組織的形成。FZP上游約5 kb處的18 bp串聯(lián)重復序列抑制其表達,延長了穗分枝歷時從而增加每穗穎花數(shù),增產(chǎn)約15%。An-1編碼bHLH蛋白,調(diào)控細胞分裂,正調(diào)控芒長和粒長,負調(diào)控每穗穎花數(shù)。An-2/ LABA1編碼LOG-like蛋白6,催化CK合成的最后一步反應(yīng),通過促進細胞分裂增加芒長,同時降低每穗穎花數(shù)和分蘗數(shù)。GAD1也是一個正調(diào)控芒長和粒長、負調(diào)控每穗穎花數(shù)的基因。另外,也發(fā)現(xiàn)了正調(diào)控每穗穎花數(shù)基因,如富含亮氨酸重復受體樣激酶基因LRK1,過表達該基因可以增加每穗穎花數(shù)。Spr3最初由林鴻宣教授課題組定位在4.6 kb區(qū)間內(nèi),但是這個區(qū)間并沒有編碼基因,通過近等基因系比較發(fā)現(xiàn),SG-64等位基因具有增加每穗穎花數(shù)的作用;而日本Ishii等將該基因定位在9.3 kb區(qū)間,OsLG1位于定位區(qū)間下游,通過互補測驗驗證了9.3 kb的DNA片段具有上調(diào)OsLG1的作用,Spr3/OsLG1的克隆證實遠端調(diào)控對基因功能起到重要的作用。LF1是一個功能獲得性突變體,穎花兩側(cè)護穎發(fā)育成側(cè)生小花,形成簇生小花,從而對每穗穎花數(shù)具有重要的調(diào)控作用。
3.4 粒形發(fā)育的分子機制
GS3是粒長的負調(diào)控因子,編碼G蛋白γ亞基,并且與G蛋白的其他亞基以及OsMADS1共同作用,調(diào)控水稻粒型。GW2負調(diào)控粒寬,編碼一個功能未知的RING-type蛋白,具有E3泛素連接酶活性,可能參與泛素蛋白酶體對蛋白的降解途徑。GIF1編碼細胞壁蔗糖酶,影響水稻灌漿,表達模式對其功能的發(fā)揮具有重要的影響,栽培稻中GIF1自身啟動子驅(qū)動GIF1的過量表達能增加粒重。
BRs是重要的植物激素,其合成和信號轉(zhuǎn)導途徑對水稻粒形具有調(diào)控作用。GS5編碼一個絲氨酸羧肽酶,GS5-1能增強BRs的信號,正調(diào)控種子大小。GW5/qSW5 (grain width 5/seed width 5)是一個控制粒寬的主效基因,前期研究認為GW5編碼一個核定位蛋白,并且1212 bp的缺失增加粒寬;隨后,進一步研究發(fā)現(xiàn)1212 bp的缺失導致了其5 kb下游處的鈣調(diào)素結(jié)合蛋白(GW5)表達量的降低,增加了粒寬。GW5定位在細胞膜上,直接與GSK2互作并抑制GSK2的激酶活性,激活BRs信號。轉(zhuǎn)錄調(diào)控因子GRF家族的GL2/GS2/PT2編碼OsGRF4,直接與BRs的負調(diào)控因子GSK2互作,抑制GSK2的轉(zhuǎn)錄激活活性,從而上調(diào)受BRs信號誘導的基因表達,激活BRs的響應(yīng)。qGL3.1/OsPPKL1編碼PPKL家族的絲氨酸/蘇氨酸磷酸酶OsPPKL1,也可能參與BRs的轉(zhuǎn)導過程。TGW6編碼IAA(indole-3-acetic acid)葡萄糖水解酶,在胚乳發(fā)育過程中對保持生長素的穩(wěn)態(tài)起著重要的作用,不僅直接控制胚乳大小,而且間接參與了同化物從源到庫的運輸及分配。
粒寬QTL GW8編碼SPL家族蛋白OsSPL16,通過增加細胞數(shù)目增加粒寬和粒重。粒長和粒寬QTL GL7/ GW7編碼與擬南芥中LONGIFOLIA蛋白的同源蛋白。GL7位點上17.1 kb片段的串聯(lián)重復,引起GL7的上調(diào)表達,從而增加水稻粒長。GW8中SBP 結(jié)構(gòu)域直接結(jié)合在GW7的啟動子,抑制GW7基因的表達,從而粒長變短。粒長粒重基因GLW7編碼SPL家族的轉(zhuǎn)錄因子OsSPL13,正調(diào)控穎殼的細胞大小,具有增加粒長和產(chǎn)量的作用。
3.5 株型的分子機制
自20世紀60年代以來,“綠色革命基因”sd1 (semi-dwarf 1)在水稻矮化育種中得到了廣泛的應(yīng)用,但是直到2002年sd1才被克隆。SD1編碼GA20氧化酶蛋白OsGA20ox2,催化GA53~GA44~GA19~ GA20的反應(yīng),活性的GA3處理sd1幼苗可以恢復到野生型表型。通過對矮桿品種中sd1的比較測序,發(fā)現(xiàn)sd1的突變主要有7種等位基因型,分別為IR8的383 bp缺失等位基因型、93-11外顯子提前終止等位基因型、矮腳南特中第一個外顯子中2 bp缺失等位基因型,以及4種SD1氨基酸改變的等位基因型,它們的廣泛利用推動了矮稈育種的進程。Sd1不僅在“第一次綠色革命”中得到選擇應(yīng)用,而且在更早的粳稻馴化過程中就被選擇;粳稻中兩個功能性的FNPs(SD1-EQ)導致GA20ox2酶活性降低,內(nèi)源GA含量降低,最終株高降低,而秈稻和野生稻攜帶了強功能型的SD1(SD1-GR),內(nèi)源GA含量升高,株高增加。
最重要的理想株型基因IPA1編碼OsSPL14,是OsmiRNA156的直接靶標。IPA1的一個點突變阻斷了OsmiRNA156介導的OsSPL14調(diào)控,使分蘗減少、穗粒數(shù)和千粒重增加,同時莖稈變得粗壯,抗倒伏能力增強,產(chǎn)量提高。在營養(yǎng)生長期,OsSPL14控制水稻分蘗;在生殖生長期,OsSPL14高表達促進了穗分支。IPA1可以直接結(jié)合在OsTB1的啟動子上,負調(diào)控水稻分蘗發(fā)生,正調(diào)控DEP1調(diào)節(jié)水稻株高和穗長。研究還發(fā)現(xiàn),超級稻甬優(yōu)12的IPA1等位基因上游具有3個3137 bp的串聯(lián)重復,使啟動子區(qū)域甲基化程度降低以及染色質(zhì)結(jié)構(gòu)松散,導致ipa1-2D的表達量上升,分蘗數(shù)變少,而穗變大,所以,適度的IPA1表達量能具有最高的產(chǎn)量潛力。
分蘗角是株型的重要性狀,普通野生稻具有匍匐生長的特性,分蘗角很大,而栽培稻表現(xiàn)為直立生長,分蘗角較小。PROG1是一個重要的控制匍匐生長習性的馴化基因,編碼Cys2-His2鋅指蛋白轉(zhuǎn)錄因子。在野生稻到栽培稻的馴化過程中,PROG1基因功能喪失,株型得到改良,每穗粒數(shù)增加,產(chǎn)量提高。TAC1是分蘗角主效QTL,編碼一個禾本科特有的蛋白,其內(nèi)含子中SNP的突變導致了tac1表達量的降低,產(chǎn)生了緊湊的株型。
3.6 開花與水稻產(chǎn)量的聯(lián)動機制
開花期不僅是水稻地域適應(yīng)性的重要性狀,在一定的范圍內(nèi),開花期與產(chǎn)量正相關(guān)。Ghd7編碼一個CCT結(jié)構(gòu)蛋白,Ghd8/DTH8編碼一個CCAAT盒結(jié)合蛋白亞基的OsHAP3,Ghd7.1/DTH7編碼一個PRR蛋白,它們都對光照長度敏感,在長日照條件下,通過抑制Ehd1、RFT1和Hd3a延遲抽穗,同時增加株高和每穗穎花數(shù)。抽穗期基因Ghd7、Ghd8、Ghd7.1和Hd1功能的減弱或缺失促成了水稻從低緯度地區(qū)向高緯度地區(qū)擴散。在長日照條件下,Hd1和Ghd7互作形成Ghd7-Hd1的蛋白復合體,特異性地結(jié)合在Ehd1的順式作用元件上,抑制Ehd1的表達延遲抽穗。在長日照條件下,Hd1也可以直接與Ghd8互作,形成Ghd8-Hd1復合體,抑制Hd3a的轉(zhuǎn)錄,從而延遲抽穗。在短日照條件下,Ghd8的轉(zhuǎn)錄得不到積累,不能形成Ghd8-Hd1復合體,而Hd1可以激活Hd3a的表達促進抽穗。
3.7 水稻重要利用價值的抗逆新基因和營養(yǎng)高效吸收基因發(fā)掘
挖掘具有抗生物逆境和非生物逆境的基因并闡明作用機理,對培育高產(chǎn)穩(wěn)產(chǎn)水稻品種具有重大意義。近20年來,一大批水稻抗病基因被克隆,而這些基因具有不同的作用方式,包括作用于病原識別、信號轉(zhuǎn)導、下游防御相關(guān)蛋白以及不同信號之間的相互作用。最近幾年抗蟲基因克隆也取得突破性進展,多個抗褐飛虱基因被克隆,如Bph3、Bph14/Qbp1、BPH18和Bph9等,為抗褐飛虱水稻品種的培育提供了優(yōu)異的基因資源。
水稻優(yōu)良品種應(yīng)具備抗非生物逆境如高溫和冷害等特性。水稻耐寒基因COLD1編碼G蛋白信號的調(diào)控因子,其與RGA1互作,激活Ca2+通道感受低溫,并加速G蛋白GTPase活性。超表達COLD1jap能增強水稻耐寒性。耐冷QTLCTB4a編碼富含亮氨酸受體蛋白激酶;上調(diào)CTB4a會增強ATP合酶活性,提高ATP含量,在低溫條件下提高結(jié)實率和產(chǎn)量。耐高溫QTL TT1編碼26S蛋白酶體α2亞基,參與泛素化蛋白降解途徑,非洲栽培稻中的等位基因具有耐高溫;高溫誘導TT1表達,可以降解有毒蛋白以及維持高溫應(yīng)答過程,從而保護細胞免受損傷。
養(yǎng)份高效利用是綠色水稻品種的重要特性。DEP1/qNGR9編碼G蛋白γ亞基,除了控制直立穗的性狀外,還影響水稻氮利用效率,DEP1能與Gα (RGA1)和Gβ (RGB1)亞基互作,共同調(diào)控氮信號。最近有研究報道GRF4可以結(jié)合在控制氮代謝、光合作用、蔗糖代謝、蔗糖轉(zhuǎn)運和細胞分裂等基因的啟動子上,上調(diào)基因的表達,促進氮素同化、固碳作用以及水稻的生長;而SLR1 (slender rice 1)編碼DELLA蛋白,活性GAs可以促進SLR1的降解,SLR1可以與GRF4拮抗,并對GRF4的功能起抑制作用,SLR1積累不僅造成矮化,還降低了氮肥使用效率。因此,可以通過增加GRF4的表達,提高含有半矮桿基因sd1品種的氮利用效率,從而進一步提高產(chǎn)量。PSTOL1/Pup1編碼Pup1特異性蛋白激酶,超表達PSTOL1能在磷缺乏條件下顯著增加產(chǎn)量。PSTOL1能增強早期根的發(fā)育,使植株獲得更多的磷和其他營養(yǎng)元素。
3.8 中國在水稻基礎(chǔ)研究中的地位和貢獻
上述大部分進展是中國科學家完成的。在NCBI中以rice為關(guān)鍵詞,提取2003年~2018年7月間發(fā)表的所有與水稻相關(guān)的論文,選取Nature Index所包含雜志中植物學相關(guān)的雜志,共篩選出1286篇論文,其中通訊作者單位為中國的有307篇(圖3)。世界每年有60~100篇高水平水稻論文發(fā)表,中國發(fā)表論文數(shù)量在其中所占比例越來越高,從2003年占比12%到2018年7月占比43%。這表明,我國在水稻領(lǐng)域的基礎(chǔ)研究突飛猛進,已經(jīng)處于國際領(lǐng)先水平,中國的重要論文貢獻份額近5年穩(wěn)定在30%左右。
4 水稻育種的發(fā)展方向
4.1 雜交育種與標記輔助選擇技術(shù)緊密結(jié)合
常規(guī)育種、標記輔助選擇育種和轉(zhuǎn)基因育種在品種選育中都表現(xiàn)出各自的優(yōu)勢。但是,當前標記輔助選擇大多是獨立于雜交育種過程,只是對雜交育種等方法選育出的品種的個別缺陷性狀進行改良,改良后新品種的基因組結(jié)構(gòu)與原品種變化很小。轉(zhuǎn)基因育種與標記輔助選擇育種一樣,也只是對現(xiàn)有品種的個別性狀加以改良。從育種的效率和效果看,毋庸置疑,水稻雜交育種方法依舊是主流方法。但是,雜交育種一定要在選育過程中與現(xiàn)代育種技術(shù)相結(jié)合,避免過去僅限于大田的純表型選擇,在雜交育種完成后再開展不良性狀的改良。因此,在開展雜交育種之前,一定要先明確育種目標,首先要篩選符合育種目標的材料作為親本,再明確親本中調(diào)控目標性狀的主要基因是否有功能或者功能強弱。這樣,在雜交育種的低世代材料中根據(jù)農(nóng)藝性狀選擇一些優(yōu)良單株,再對這些單株進行標記輔助選擇,從中選擇攜帶無法目測的優(yōu)良目標基因的單株,進入下一個世代,在雜交育種過程中完成標記輔助選擇,使新品種具有育種目標性狀,沒有明顯缺陷,省去輔助選擇對某個性狀修修補補的必要。
4.2 滿足消費者需要,培育口感好的品種
隨著生活水平的提高,人們對稻米品質(zhì)有了更高的追求。除了外觀品質(zhì)和營養(yǎng)品質(zhì)外,口感應(yīng)該成為品質(zhì)的重要指標。口感與水稻中直鏈和支鏈淀粉的組成比例和蛋白含量密切相關(guān),但是影響口感的遺傳基礎(chǔ)還不清晰,需要加強遺傳學研究。
4.3 滿足輕簡栽培需要,培育適合新的耕作制度的品種
直播和機械化等輕簡栽培技術(shù)大大降低了勞動力成本,已成為水稻生產(chǎn)的主流技術(shù)。然而,輕簡栽培出現(xiàn)新的生產(chǎn)問題:直播稻易倒伏,增加機械化收獲難度。因此,培育莖稈粗壯、增強抗倒伏特性的品種有利于輕簡栽培技術(shù)技術(shù)推廣。另外,當前耕作制度多樣化,從兩季稻變一季稻,或者一種兩收的再生稻。特別是張啟發(fā)先生提出的長江中下游稻區(qū)“雙水雙綠”稻蝦共生的種植/養(yǎng)殖模式,具有較高的經(jīng)濟效益。為滿足稻蝦連作的需求,應(yīng)該培育生育期適宜、莖稈粗壯、抗倒伏和抗病蟲害的水稻品種。
4.4 滿足生態(tài)環(huán)保需要,培育具有特色健康品質(zhì)的綠色新品種
培育抗病蟲害的綠色品種是生產(chǎn)健康稻米的基本條件。水稻中一系列的抗稻瘟病和白葉枯病基因,以及抗飛虱基因得到定位或克隆。但是還沒有找到有效抗稻曲病和抗螟蟲的基因,今后需要在這些方面力爭突破。土壤重金屬污染特別是鎘污染尤其嚴重,對食品安全和人類健康有重要的影響。2010年,Ueno等克隆出一個控制水稻鎘積累的基因OsHMA3,超表達低鎘積累的等位基因可以選擇性的降低種子中鎘的積累,而對其他的微量元素沒有影響。選育種子中鎘或其他重金屬含量低的品種,解決“重金屬米”的安全隱患。另外,應(yīng)該重視培育富含微量元素如富硒水稻品種,重視培育滿足特殊人群需要的水稻品種,如適合糖尿病患者食用的抗性淀粉高的稻米等。
總之,水稻育種要緊跟耕作制度變化的步伐,培育適合新耕作方法的新品種。一方面要利用基因組育種技術(shù)和基因編輯技術(shù),加快水稻功能基因組研究成果向育種應(yīng)用的轉(zhuǎn)化,另一方面要重視發(fā)掘新的重要基因,為設(shè)計育種提供元件!培育高產(chǎn)優(yōu)質(zhì)的綠色超級稻品種是人心所向,大勢所趨。
作者單位系華中農(nóng)業(yè)大學作物遺傳改良國家重點實驗室